My research interests lie broadly in understanding the evolution of novel floral traits. For my dissertation, I am working to understand the ecological function and genetic basis of personate flowers in Penstemon wildflowers. Personate flowers are characterized by an upward bulge in the lower lip of the corolla tube that ultimately seals off the floral passageway (see P. hirsutus). This floral trait has evolved at least two times in the genus Penstemon, but no previous research has focused on understanding why or how this trait evolved.


Several of my experiments use the personate Penstemon hirsutus (left) and open-tubed Penstemon smallii (right) as my focal species because they represent contrasting extremes of floral morphology.
Below, you will find information regarding several of my dissertation projects!
What is the morphological basis of personate flowers?
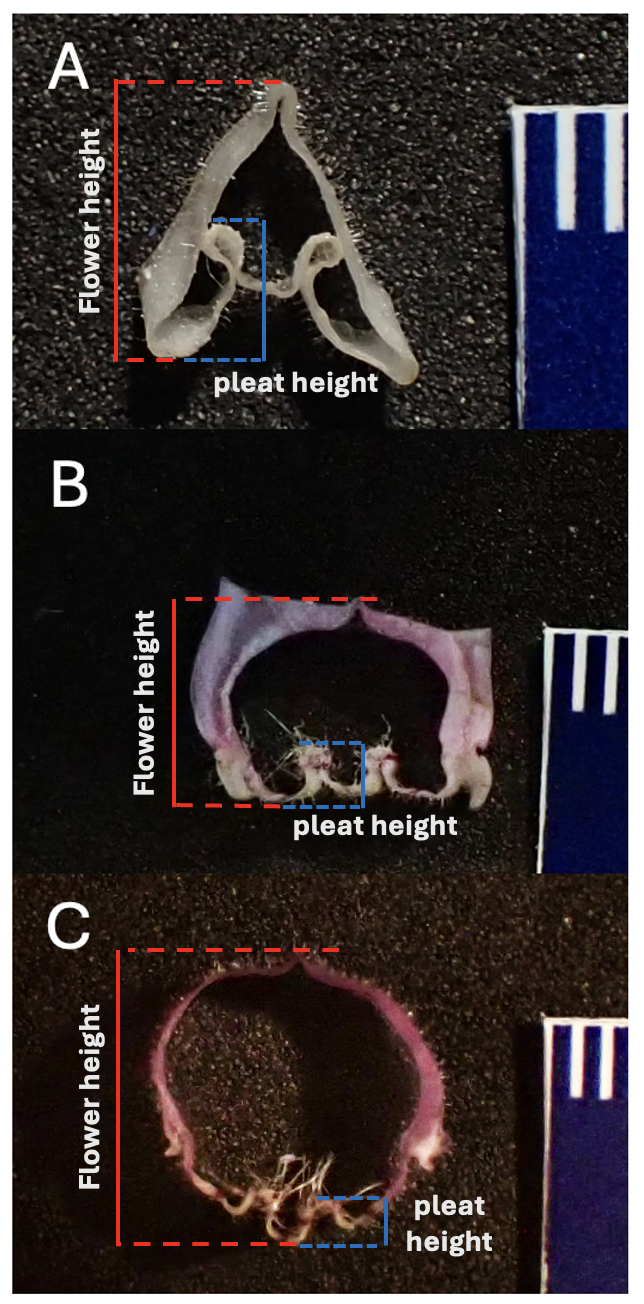
Understanding morphological characteristics that differentiate personate flowers from open-tubed flowers is a necessary component for my research. I have used several different methods including size-standardized photos, microCT scanning, and geometric morphometrics. To date, the best method for quantifying a flower’s degree of personate-ness is by taking photographs of cross-sections sliced through the flower (see right). With these photos, I measure the height of the “pleat” that pushes into the flower and the total flower height. I compare these measures to each other for my “Pleat Depth” metric.

Above: Three dimensional rendering of personate flowered P. hirsutus from MicroCT scans.
Right: Cross-section cuts through flowers that clearly show the difference in flower shape between personate (top), tubular (middle), and open (bottom)
How many times have personate flowers evolved in the Eastern North American Penstemons?
The presence of three personate species in Penstemon’s Eastern United States subsection (subsect. Penstemon) provides an excellent opportunity to determine how many times this novel floral trait has evolved. Are all three personate Penstemon species each other’s closest relative? Or is there a deviation from what we would expect based on the species’ shared phenotype? If there are multiple origins of personate flowers, is this due to a simple genetic architecture underlying personate flowers? Contrarily, perhaps adaptive introgression has moved this trait between lineages! To understand the evolutionary history of personate flowers I generated whole-genome resequencing data (Novaseq) for all diploid species in Penstemon subsect. Penstemon, inferred species relationships, and calculated introgression metrics.
You can read about these results and more in the paper!



I am using a combination of pollinator observations and a mating system study to answer this question!
The pollinator observations provide me with the opportunity to determine if personate flowers are filtering pollinators in any way. My initial hypothesis was that, similar to previous work in Snapdragon, Penstemon’s personate flowers would filter bees based on size. However, results from my 2022 field season does not support this hypothesis. Instead, I found that both large (i.e., Bombus) and small (i.e., Osmia and Ceratina) bees visit both floral types.
The mating system study is driven by my observation that the upward bulge in the lower lip of personate flowers causes the anthers and stigma to be closer to each other (compared to their positions in the open-tubed flowers). My hypothesis is that the decreased distance between reproductive structures makes personate flowers better adapted to selfing. To test this hypothesis, I assign both open-tubed and personate flowers to different treatment groups and then quantify seed set to determine the fitness of each group.
What is the ecological function of personate flowers?


What is the genetic basis of personate flowers?
Floral trait variation has proven to be extremely useful for understanding the genetic mechanisms underlying adaptation – a major goal of evolutionary genetics.
To determine how personate flowers evolved I conducted a Quantitative Trait Locus (or QTL) mapping analysis. Broadly, this analysis identifies significant correlations between phenotype and genotype data to inform on the number and location of loci responsible for a trait. My QTL analysis identified two unlinked loci accounting for 51% of the variation in Penstemon’s personate flowers.

I designed an RNA-Seq experiment to detect Differentially Expressed (DE) genes between focal Penstemon species in ventral tissue across multiple developmental stages. More specifically, I collected flower buds from the parents of my QTL cross, P. hirsutus and P. smallii, at two different developmental time points and dissected the buds to separate the dorsal and ventral floral tissues. Both tissue types, stages and species were sequenced separately and used in subsequent RNA-Seq analyses to identify a handful of candidate genes responsible for personate flowers.
Which genes are responsible for personate flower production?
